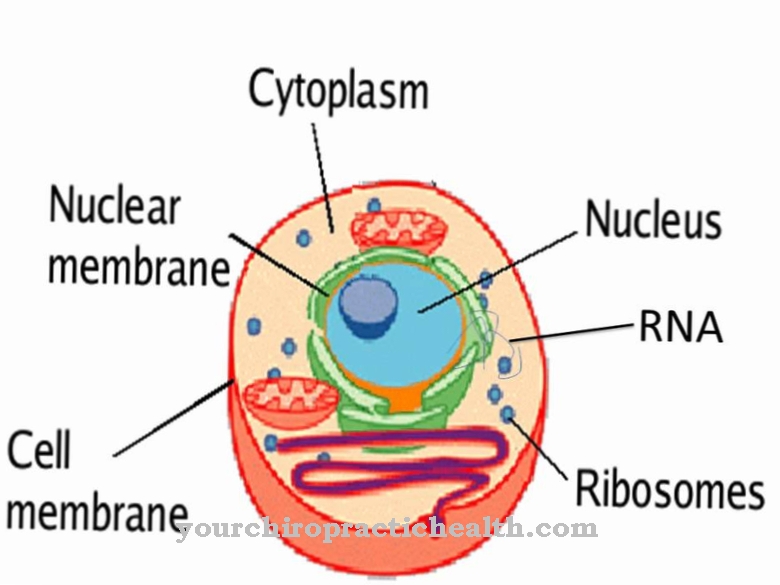
Histoni ir daļa no šūnu kodoliem. To klātbūtne ir atšķirības pazīme starp vienšūnu organismiem (baktērijām) un daudzšūnu organismiem (cilvēkiem, dzīvniekiem vai augiem). Ļoti nedaudzos baktēriju celmos ir olbaltumvielas, kas ir līdzīgas histoniem. Evolūcija ir radījusi histonus, lai labāk un efektīvāk augstāko dzīvo lietu šūnās varētu ievietot ļoti garo DNS ķēdi, kas pazīstama arī kā ģenētiskais materiāls. Tā kā, ja cilvēka genoms tiktu izjaukts, tas būtu apmēram 1-2 m garš, atkarībā no šūnas stadijas, kurā atrodas šūna.
Kas ir histones?
Augstāk attīstītos organismos histoni rodas šūnu kodolos, un tiem ir liels pozitīvi lādētu aminoskābju (īpaši lizīna un arginīna) īpatsvars. Histonu olbaltumvielas ir sadalītas piecās galvenajās grupās - H1, H2A, H2B, H3 un H4. Četru grupu H2A, H2B, H3 un H4 aminoskābju secības gandrīz neatšķiras starp dažādām dzīvām būtnēm, turpretim H1, kas savieno histonu, ir vairāk atšķirību. Putnu sarkanās asins šūnas, kas satur kodolu, H1 pat ir pilnībā aizstāts ar citu galveno histonu grupu - H5.
Lielā sekvenču līdzība lielākajā daļā histonu olbaltumvielu nozīmē, ka lielākajā daļā organismu DNS "iesaiņošana" notiek vienādi un iegūtā trīsdimensiju struktūra ir vienlīdz efektīva histonu funkcijai. Evolūcijas gaitā histonu attīstībai ir jānotiek ļoti agri, un tai jābūt uzturētai pat pirms zīdītāju vai cilvēku parādīšanās.
Anatomija un struktūra
Tiklīdz šūnā no atsevišķām bāzēm (ko sauc par nukleotīdiem) veidojas jauna DNS ķēde, tā ir "jāiesaiņo". Šim nolūkam histone olbaltumvielas dimerizējas, kas pēc tam katrs veido divus tetramerus. Visbeidzot, histona kodols sastāv no diviem tetrameriem, histona oktamera, ap kuru DNS šķipsna tiek ietīta un daļēji iespiesta. Tādējādi histona oktamers atrodas trīsdimensiju struktūrā savītas DNS virknes ietvaros.
Astoņi histonu proteīni ar DNS ap tiem veido visu nukleosomas kompleksu. DNS laukumu starp divām nukleosomām sauc par saistītāj DNS, un tas satur apmēram 20-80 nukleotīdus. Linkera DNS ir atbildīga par DNS "ievadīšanu" un "aiziešanu" no histona oktamera. Tādējādi nukleosoma sastāv no aptuveni 146 nukleotīdiem, linkera DNS daļas un astoņiem histona proteīniem, tā, lai 146 nukleotīdi apvītot histona oktameru apvidu 1,65 reizes.
Turklāt katra nukleosoma ir saistīta ar H1 molekulu tā, ka DNS ieejas un izejas punktus tur kopā savienojošais histons un palielinās DNS kompakcija. Nukleosomas diametrs ir aptuveni 10–30 nm. Daudzas nukleosomas veido hromatīnu - garu DNS-histona ķēdi, kas zem elektronu mikroskopa izskatās kā pērļu virtene. Nukleosomas ir "pērles", kuras ieskauj vai savieno virknei līdzīga DNS.
Vairāki nehistone proteīni atbalsta atsevišķu nukleosomu vai visa hromatīna veidošanos, kas galu galā veido atsevišķas hromosomas, ja šūna dalās. Hromosomas ir maksimālais hromatīna saspiešanas veids, un to var atpazīt ar gaismas mikroskopiju šūnas kodola dalīšanas laikā.
Funkcija un uzdevumi
Kā minēts iepriekš, histoni ir pamata olbaltumvielas ar pozitīvu lādiņu, tāpēc tie mijiedarbojas ar negatīvi lādētu DNS, izmantojot elektrostatisko pievilcību. DNS "aptin" ap histona oktimēriem, lai DNS kļūtu kompakta un iekļautos katras šūnas kodolā. H1 funkcija ir saspiest augstāka līmeņa hromatīna struktūru un lielākoties novērš transkripciju un tādējādi translāciju, t.i., šīs DNS daļas translāciju olbaltumvielās caur mRNS.
Atkarībā no tā, vai šūna "atpūšas" (starpfāzes) vai dalās, hromatīns ir mazāk vai spēcīgāk kondensēts, t.i., iesaiņots. Starpfāzē lielas hromatīna daļas ir mazāk kondensētas, tāpēc tās var pārrakstīt mRNS, t.i., nolasīt un vēlāk pārveidot olbaltumvielās. Histoni regulē atsevišķu gēnu aktivitāti to tuvumā un ļauj transkripcijai un mRNS dzīslu izveidošanai.
Kad šūna sāk dalīties, DNS netiek pārveidota olbaltumvielās, bet vienmērīgi sadalīta starp divām izveidotajām meitas šūnām. Tāpēc hromatīns ir stipri kondensēts un papildus stabilizēts ar histoniem. Hromosomas kļūst redzamas un ar daudzu citu nehistoņu olbaltumvielu palīdzību var tikt sadalītas jaunattīstības šūnās.
Slimības
Histoniem ir būtiska nozīme jaunas dzīvās būtnes radīšanā. Ja histonu gēnu mutāciju dēļ nevar veidoties viens vai vairāki histona proteīni, šis organisms nav dzīvotspējīgs un tā priekšlaicīga turpmāka attīstība tiek apturēta. Tas galvenokārt ir saistīts ar histonu augsto secību.
Tomēr kādu laiku ir zināms, ka bērniem un pieaugušajiem ar dažādiem ļaundabīgiem smadzeņu audzējiem mutācijas var rasties dažādos audzēja šūnu histonu gēnos. Histonu gēnu mutācijas ir aprakstītas galvenokārt tā sauktajās gliomās. Šajos audzējos tika atklātas arī iegarenas hromosomu astes. Šīs hromosomu gala sadaļas, ko sauc par telomeriem, parasti ir atbildīgas par hromosomu ilgmūžību. Šajā kontekstā šķiet, ka iegarenie telomēri audzējos ar histona mutācijām dod šīm deģenerētajām šūnām izdzīvošanas priekšrocības.
Pa to laiku ir zināmi citi vēža veidi, kuriem ir mutācijas dažādos histonu gēnos un tādējādi rodas mutētas histona olbaltumvielas, kuras nepilda vai tikai slikti pilda savus regulatīvos uzdevumus. Šie atradumi pašlaik tiek izmantoti, lai izstrādātu terapijas formas īpaši ļaundabīgiem un agresīviem audzējiem.


.jpg)


.jpg)



.jpg)

.jpg)
.jpg)