No Taukskābju sadalīšana kalpo enerģijas ģenerēšanai šūnās un notiek caur tā saucamo beta oksidāciju. Beta oksidācijas laikā veidojas acetilkoenzīms A, kas tālāk tiek sadalīts oglekļa dioksīdā un ūdenī vai atkal ievadīts citronskābes ciklā. Traucējumi taukskābju sadalījumā var izraisīt nopietnas slimības.
Kas ir taukskābju sadalījums?

Papildus glikozes sadalīšanai organismā, taukskābju sadalīšanās ir svarīgs metabolisma process enerģijas ģenerēšanai šūnā.
Taukskābes tiek sadalītas mitohondrijos. Noārdīšanās notiek, izmantojot tā saukto beta oksidāciju. Termins "beta" radās no tā, ka taukskābju molekulas trešais oglekļa atoms (beta oglekļa atoms) oksidējas.
Oksidācijas cikla beigās divi oglekļa atomi tiek sadalīti aktivētās etiķskābes (acetilkoenzīma A) veidā. Tā kā taukskābju sadalīšanai nepieciešami vairāki oksidācijas cikli, process agrāk bija pazīstams arī kā taukskābju spirāle.
Acetilkoenzīms mitohondrijos tiek sadalīts tālāk ketonu ķermeņos vai oglekļa dioksīdā un ūdenī. Pēc mitohondrija nonākšanas citoplazmā tas tiek ievadīts citronskābes ciklā.
Taukskābju sadalīšanās laikā tiek saražots vairāk enerģijas nekā sadedzinot glikozi.
Funkcija un uzdevums
Taukskābju sadalīšana notiek vairākos reakcijas posmos un notiek mitohondrijos. Pirmkārt, taukskābju molekulas atrodas šūnas citosolā.
Tās ir inertas molekulas, kuras vispirms jāaktivizē sadalīšanai un jāpārvadā mitohondrijos. Lai aktivizētu taukskābi, koenzīms A tiek pārnests, veidojot acil-CoA. Pirmkārt, ATP tiek sadalīts pirofosfātā un AMP. Pēc tam AMP izmanto, lai izveidotu acil-AMP (acil-adenilātu).
Pēc AMP sadalīšanas taukskābes var esterizēt ar koenzīmu A, lai veidotu acil-CoA. Pēc tam ar fermenta karnitīna aciltransferāzes I palīdzību karnitīns tiek pārnests uz aktivēto taukskābi.
Šo kompleksu ar karnitīna-acilkarnitīna transportētāju (CACT) transportē mitohondrijā (mitohondriju matricā). Tur karnitīns atkal tiek sadalīts un atkal tiek pārnests koenzīms A. Karnitīns tiek novirzīts no matricas, un mitohondrijā ir pieejama acil-CoA faktiskajai beta oksidācijai.
Faktiskā beta oksidācija notiek četrās reakcijas stadijās. Klasiskās oksidācijas pakāpes notiek ar pāra numurētām piesātinātajām taukskābēm. Ja tiek sadalītas nepāra vai nepiesātinātās taukskābes, sākuma molekula vispirms jāsagatavo beta oksidēšanai, izmantojot turpmākas reakcijas.
Pāra numurētu piesātināto taukskābju acil-CoA tiek oksidēts pirmajā reakcijas posmā ar enzīma acil-CoA dehidrogenāzes palīdzību. Tas rada divkāršu saiti starp otro un trešo oglekļa atomu trans stāvoklī. Turklāt FAD tiek pārveidots par FADH2.
Parasti nepiesātināto taukskābju divkāršās saites atrodas cis stāvoklī, bet nākamais taukskābju sadalīšanās reakcijas solis var notikt tikai ar divkāršo saiti trans stāvoklī.
Otrajā reakcijas posmā enzīma enoil-CoA hidratāze pievieno ūdens molekulu beta oglekļa atomam, veidojot hidroksilgrupu. Tad tā sauktā L-3-hidroksi-acil-CoA dehidrogenāze oksidē beta oglekļa atomu keto grupai. Veidojas 3-ketoacil-CoA.
Pēdējā reakcijas posmā papildu koenzīms A saistās ar beta-oglekļa atomu. Acetil-CoA (aktivētā etiķskābe) tiek sadalīta, un paliek acil-CoA, kas ir par diviem oglekļa atomiem īsāks. Šī īsākā atlikušā molekula iziet cauri nākamajam reakcijas ciklam līdz turpmākai acetil-CoA šķelšanai.
Process turpinās, līdz visa molekula tiek sadalīta aktivētā etiķskābē. Beta oksidācijas apgrieztais process arī būtu teorētiski iespējams, taču dabā tas nenotiek.
Taukskābju sintēzei ir atšķirīgs reakcijas mehānisms. Mitohondrijā acetil-CoA tiek sadalīta tālāk oglekļa dioksīdā un ūdenī vai ketonu ķermeņos, atbrīvojot enerģiju. Nepāra numuru taukskābju gadījumā propionil-CoA ar trim oglekļa atomiem paliek beigu daļā. Šī molekula tiek sadalīta atšķirīgā veidā.
Sadalot nepiesātinātās taukskābes, ar īpašām izomerāzēm divkāršās saites no cis tiek pārveidotas par trans-konfigurācijām.
Slimības un kaites
Taukskābju sadalīšanās traucējumi ir reti, taču tie var izraisīt nopietnas veselības problēmas. Gandrīz vienmēr tās ir ģenētiskas slimības.
Gandrīz katram attiecīgajam enzīmam, kas iesaistīts taukskābju sadalīšanā, ir atbilstoša gēna mutācija. Piemēram, MCAD enzīma deficīts rodas no gēna mutācijas, kas tiek mantota autosomāli recesīvā veidā. MCAD ir atbildīgs par vidējās ķēdes taukskābju sadalīšanu. Simptomi ir hipoglikēmija (zems cukura līmenis asinīs), krampji un biežas komomas stāvokļi. Tā kā taukskābes šeit nevar izmantot enerģijas iegūšanai, paaugstināts glikozes līmenis tiek sadedzināts. Tas izraisa hipoglikēmiju un komas risku.
Tā kā ķermenis enerģijas iegūšanai vienmēr ir jāpiegādā ar glikozi, ilgstoši nedrīkst atturēties no pārtikas. Ja nepieciešams, akūtas krīzes gadījumā jāveic lielas devas glikozes infūzija.
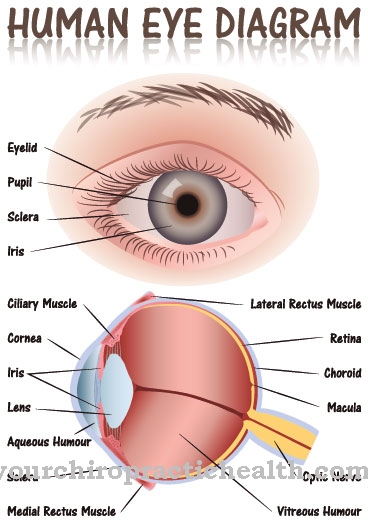
Turklāt visas miopātijas ir raksturīgas mitohondriju taukskābju sabrukšanas traucējumiem. Tas noved pie muskuļu vājuma, aknu metabolisma traucējumiem un hipoglikēmiskiem stāvokļiem. Līdz 70 procentiem skarto cilvēku dzīves laikā kļūst akli.
Nopietnas slimības rodas arī tad, ja tiek traucēta pārmērīgi garu taukskābju sadalīšanās. Šīs ļoti garās ķēdes taukskābes netiek sadalītas mitohondrijos, bet peroksisomās. Ferments ALDP ir atbildīgs par ievadīšanu peroksisomās. Tomēr, ja ALDP ir nepilnīgs, garo taukskābju molekulas uzkrājas citoplazmā un tādējādi noved pie smagiem vielmaiņas traucējumiem. Uzbrūk arī nervu šūnas un smadzeņu baltā viela. Šāda veida taukskābju sabrukšanas traucējumi izraisa neiroloģiskus simptomus, piemēram, līdzsvara traucējumus, nejutīgumu, krampjus un zemādas virsnieru dziedzerus.




.jpg)






.jpg)